Mini-Review: F4/80 Antibody
F4/80: The Marker and the Antibody
F4/80 is a well-characterized and extensively referenced membrane protein and is the best-known mature mouse macrophage and microglial marker.
The anti-F4/80 antibody clone CI:A3-1 is the original monoclonal produced against the F4/80 antigen in Siamon Gordon’s laboratory at the University of Oxford (Gordon and Austyn 1981). Today, Bio-Rad is the only commercial manufacturer of the original clone for this important monoclonal macrophage antibody.
Over 44 formats of F4/80 antibody are currently available, from purified and low endotoxin versions to a wide range of popular fluorescent labels. These fluorescent labels are suitable for flow cytometry and include choices such as FITC, RPE, DyLight, and Alexa Fluor dyes, as well as the exclusive tandem conjugates RPE-Alexa Fluor 647 and RPE-Alexa Fluor 750.
F4/80 is now available conjugated to Bio-Rad’s innovative range of StarBright™ Dyes, with conjugates available for the ultraviolet, violet, blue, red, and yellow laser lines. StarBright Dyes have been designed to be stable with superior brightness and narrow excitation and emission characteristics, making them ideal for inclusion in your multicolor flow cytometry panels.
Discover the world's largest collection of F4/80 antibody formats (clone Cl:A3-1), including the innovative StarBright Dyes.
Find Out MoreCharacteristics of Anti-F4/80 Antibody Clone CI:A3-1
Not only can this antibody be used to detect Kupffer cells, Langerhans cells, peritoneal macrophages, and splenic red pulp macrophages, but the anti-F4/80 antibody clone CI:A3-1 can also be used to stain microglia (macrophages of the central nervous system [CNS]), giving you the added benefit of studying neurology and the growing field of neuroimmunology (Lawson et al. 1990).
In addition to its extensive use in flow cytometry, clone Cl:A3-1 is also suitable for functional studies and is available in a low endotoxin format for this purpose. The rat anti-mouse F4/80 antibody clone CI:A3-1 is also paraffin-reactive, allowing for immunohistochemical studies of paraffin-embedded sections of brain and other tissues.
Hundreds of scientific references support the use of the antibody clone Cl:A3-1 in a wide range of applications and for a variety of experimental studies. You can peruse the associated references to see how other researchers are utilizing this antibody to achieve their research goals.
Despite intensive investigation, however, the function of the F4/80 antibody target protein is not fully understood.
F4/80: Gene and Protein
The following section contains some general information that you may need to know when working with F4/80. F4/80 is known under several alternate names: Ly71, Gpf480, TM7LN3, DD7A5-7, EGF-TM7, and EMR1.
The F4/80 gene is located on mouse chromosome 17 and encodes a polypeptide of 931 amino acids that is processed into a mature protein of 904 amino acids. Note that while the predicted mass of the protein is 98.9 kD, the F4/80 protein runs at an apparent weight of 160 kD in SDS-PAGE. The mass difference is probably accounted for by extensive glycosylation.
Two papers describe the early cloning of the gene and the characterization of the protein in 1996 and 1997, McKnight et al. and Lin et al., respectively.
The N-terminus of the F4/80 polypeptide contains seven tandem EGF-like domains and shows a high degree of homology to proteins such as fibrillin-1 and fibulin-2. The C- terminal sequence demonstrates homology to members of the TM7 superfamily, such as the G protein-coupled receptors for peptide hormones, e.g., parathyroid hormone, calcitonin, and glucagon (McKnight et al. 1996).
The later discovery of other proteins with the same domain arrangement defined a new protein family named EGF-TM7, whose members include F4/80, EMR1 (the human F4/80 homolog), and human CD97 (McKnight et al. 1998).
The presence of an adhesion and a signaling domain in the F4/80 sequence suggests that it functions as a signaling molecule upon binding with other as yet unidentified cell-surface proteins. Ligands identified for other members of the EGF-TM7 protein family support this hypothesis, making it reasonable to conclude that F4/80 could have similar adhesion and signaling properties (McKnight et al. 1998).
F4/80: Functions
To support you in studying the function of F4/80, Bio-Rad provides a vast selection of antibodies with performance guarantees in key immunological applications including flow cytometry, immunohistochemistry, western blotting, and immunofluorescence. Additionally, F4/80 knockout mice can be used, which appear vital, healthy, and fertile.
Previously, Schaller et al. (2002). found that the development and antimicrobial activity of macrophages in these animals also seems to be normal.
However, Warschkau and Kiderlen (1999) have shown that administration of the anti-F4/80 antibody inhibits the production of cytokines (namely TNFa, IL-12, and IFN) from whole spleen cell cultures of SCID mice exposed to heat killed Listeria monocytogenes. The modulated cytokine response is believed to be dependent on the interaction of natural killer (NK) cells and macrophages in culture.
F4/80 is also expressed by eye-derived antigen-presenting cells (APCs) involved in anterior chamber-associated immune deviation (ACAID), a process that elicits tolerance for an antigen inoculated into the eye. ACAID is known to be antigen-specific (Wang et al. 2001). Lin et al. (2005) went on to demonstrate the involvement of F4/80 in ACAID, in particular in the generation of efferent T regulatory (Treg) cells that are the effectors of tolerance.
Without knowing the cognate ligand of F4/80, Lin et al. (2005) could not describe a molecular mechanism for the involvement of F4/80 in ACAID, but they hypothesized that F4/80 is implicated in the interaction between tolerogenic APCs and NKT cells in the spleen that is essential for the development of the CD8+ Treg cells seen in ACAID.
In the same report, Lin et al. (2005) showed that F4/80 is also needed for CD8+ Treg cell induction in low-dose oral tolerance. Furthermore, F4/80 is expressed on APCs in the Peyer’s patches (Makala et al. 2003) and on the majority of dendritic cells (DCs) in the CNS (Suther et al. 2003). These findings suggest that F4/80 could be involved in tolerance in systems distinct from ACAID and could be an essential part of the interaction between NK cells, macrophages, and F4/80 expressing APCs.
F4/80: Expression
To optimally design and analyze your experiments, it’s important to know what cells express F4/80 to accurately detect and distinguish between cell types. F4/80 is expressed at high levels on the surface of various macrophages: Kupffer cells, splenic red pulp macrophages, microglia, gut lamina propria, and Langerhans cells in the skin. Macrophages of the connective tissue, heart, kidney, reproductive, and neuroendocrine systems also express F4/80 (Gordon and Austyn 1981).
Although F4/80 is broadly represented in macrophages, its expression varies depending upon the state of maturation, developmental processes, and type of macrophage. This allows the further subclassification of cells in your experiments.
For example, Sprenger-Svačina et al. (2024) recently utilized an F4/80 antibody in flow cytometry to determine the origin of macrophages found in mouse models of autoimmune and traumatic peripheral nerve injury. By analyzing the expression levels of F4/80, they were able to distinguish between tissue-resident macrophages (CD11b+F4/80hi) and macrophages derived from the blood (CD11b+F4/80int).
In addition, Langerhans cells are known to downregulate F4/80 expression after antigen uptake. This downregulation precedes migration to the local lymph nodes and is in accord with the hypothesized adhesion and signaling function of the protein (Gordon and Austyn 1981).
Monocytes that circulate in the bloodstream also express F4/80 on the surface, but the level is lower than on tissue macrophages, indicating another correlation between the level of F4/80 and the adhesion properties of the corresponding mononuclear blood cell (Gordon and Austyn 1981).
Therefore, the use of F4/80 staining, alongside other cell-specific markers, can not only allow you to distinguish between cell types, but also, in some cases, to determine the activation or developmental state of the cell.
For optimal immunohistochemical staining of F4/80 (Figure 1), antigen unmasking procedures are recommended, as prolonged formalin fixation and the paraffin-embedding process can mask the F4/80 antigen. Bio-Rad provides detailed procedures for antigen retrieval in F4/80 staining of paraffin-embedded tissues.

Fig. 1. Liver tissue was fixed and then stained with MCA497 after pre-treatment with citrate buffer at pH 6.0. Brown patches represent stained macrophages.
F4/80: Microglia
The ability of clone CI:A3-1 to stain microglia is an advantageous feature of F4/80, as several other antigens, such as CD4, are downregulated once they pass through the blood-brain barrier (Perry and Gordon 1991).
F4/80+ microglia are found throughout the white and grey matter. Separate regions of the parenchyma contain F4/80+ cells varying in morphology. You can exploit this in your experiments to distinguish microglia from other macrophage populations in the choroid plexus and leptomeninges (Gordon et al. 1992). Lawson et al. (1990) were able to produce a map of the distribution and morphology of microglia in the adult mouse CNS due to F4/80 labeling.
Pow et al. (1989) found that F4/80+ microglia in the posterior pituitary selectively endocytose terminals of neuroendocrine cells containing oxytocin/vasopressin, indicating that hormonal stimulation could be a functional response of F4/80+ microglia.
Microglia are associated with a number of disorders, such as Alzheimer’s disease, multiple sclerosis, and prion diseases, which means that CI:A3-1 is a key F4/80 antibody clone to use in neurological research.
|
A
|
B
|
C
|
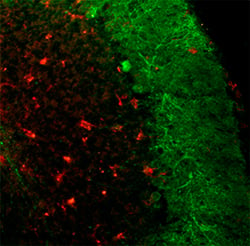
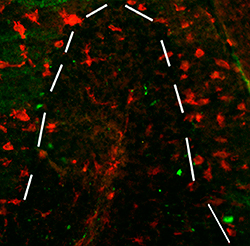
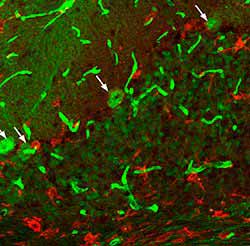
Fig. 2. These images show sections from 50-day old npc-1 mutant mice stained with anti-F4/80 antibody (red) with the aim of marking microglia. A) shows an infiltration of microglia in the granule cell layer and white matter tract. B and C) show numerous microglia marked by anti-F4/80 present throughout the cerebellum, the dashed line indicating the edge of the granule cell layer (Ko et al. 2005).
F4/80: Tumor-Associated Macrophages
Tumor-associated macrophages (TAMs) are a highly abundant cell type in the tumor microenvironment (TME) of solid cancers, the presence of which has been correlated with increased metastatic potential and poor clinical outcomes in a variety of cancer types (Chen et al. 2019; Stavrou and Constantinidou 2024; Yagi T et al. 2019).
Staining with F4/80 is one of the key methods of identifying these cells within the tumor, representing an additional emerging application of F4/80 in the field of cancer research.
TAMs have been shown to suppress antitumor activity via the induction of T-cell apoptosis and the recruitment of Tregs, and to promote tumor growth via the induction of angiogenesis (Kusmartsev and Gabrilovich 2005; Qiao et al. 2023; Wang et al. 2017).
These studies used F4/80 antibodies to study TAMs via western blotting, confocal microscopy, and flow cytometry, demonstrating the diversity of this macrophage marker in practical research. These studies could also serve as a backbone for building protocols for your own scientific goals.
Sánchez-Paulete et al. (2023) recently engineered chimeric antigen receptor (CAR) T cells to specifically target F4/80 expressing cells (F4.CAR-T cells) in a mouse model of lung cancer. Interestingly, F4.CAR-T cells were able to effectively infiltrate tumors and kill TAMs, which resulted in delayed tumor growth and significantly extended survival in mice. These data point towards the targeting of F4/80+ TAMs in cancer as a promising avenue for therapeutic potential.
Bio-Rad offers a range of antibodies, proteins, and reagents that can be used alongside the F4/80 antibody in your assays to investigate various aspects of cancer biology.
EMR1: The Human Homolog of F4/80
The human homolog of F4/80 was cloned and named EMR1 by Baud et al. (1995). EMR1 is located on chromosome 19 and its sequence shows 68% overall identity to F4/80. EMR1 also contains six EGF-like repeats and seven transmembrane segments supporting a purported function in adhesion and signaling that is in line with members of the EGF-TM7 family.
If you intend to study the human homolog EMR1 or use it in your research, there are several key differences from the mouse F4/80 antibody that you need to keep in mind. A critical divergence is the expression pattern of EMR1.
Hamann et al. (2007) studied the expression and surface abundance of EMR1 using quantitative real-time PCR and flow cytometric analysis. They observed that EMR1 is surprisingly absent on mononuclear phagocytic cells, such as monocytes, macrophages, and myeloid dendritic cells. According to their report, EMR1 expression is restricted to eosinophilic granulocytes, making it a highly specific eosinophil marker in humans rather than a macrophage marker.
While the absence of F4/80 on human monocytes and macrophages merits further investigation, the presence of F4/80 on eosinophils is not new to scientists. Murine eosinophils are known to stain with F4/80 and even Mac-1, but at a lower intensity than is typical for macrophages and monocytes (McGarry et al. 1991).
Bio-Rad offers anti-human EMR1 antibody for the study of the human equivalent of F4/80.
Related to F4/80
References
- Austyn JM and Gordon S (1981). F4/80, a monoclonal antibody directed specifically against the mouse macrophage. Eur J Immunol 11, 805–815.
- Baud V (1995). EMR1, an unusual member in the family of hormone receptors with seven transmembrane segments. Genomics 26, 334–344.
- Chen Y et al. (2019). Tumor-associated macrophages: an accomplice in solid tumor progression. J Biomed Sci 26, 78.
- Gordon S et al. (1992) Antigen markers of macrophage differentiation in murine tissues. Curr Top Microbiol Immunol 181, 1–37.
- Hamann J et al. (2007) EMR1, the human homolog of F4/80, is an eosinophil-specific receptor. Eur J Immunol 37, 2797–2802.
- Hume DA et al. (1984). The mononuclear phagocyte system of the mouse defined by immunohistochemical localization of antigen F4/80: macrophages of endocrine organs. Proc Natl Acad Sci USA 81, 4174–4177.
- Kusmartsev S and Gabrilovich DI (2005). STAT1 signaling regulates tumor-associated macrophage-mediated T cell deletion. J Immunol 174, 4880–4891.
- Ko DC et al. (2005). Cell-autonomous death of cerebellar purkinje neurons with autophagy in Niemann-Pick type C disease. PLoS Genet 1, 81–95.
- Lawson LJ et al. (1990). Heterogeneity in the distribution and morphology of microglia in the normal adult mouse brain. Neuroscience 39, 151–170.
- Lin HH et al. (1997). Identification and characterization of a seven transmembrane hormone receptor using differential display. Genomics 41, 301–308.
- Lin HH et al. (2005). The macrophage F4/80 receptor is required for the induction of antigen-specific efferent regulatory T cells in peripheral tolerance. J Exp Med 201, 1615–1625.
- Makala LHC et al. (2003). Phenotype and function of murine discrete Peyer’s patch macrophage derived - dendritic cells. J Vet Med Sci 65, 491–499.
- McGarry MP and Stewart CC (1991). Murine eosinophil granulocytes bind the murine macrophage-monocyte specific monoclonal antibody F4/80. J Leukoc Biol 50, 471–478.
- McKnight AJ et al. (1996). Molecular cloning of F4/80, a murine macrophage-restricted cell surface glycoprotein with homology to the G-protein-linked transmembrane 7 hormone receptor family. J Biol Chem 271, 486–489.
- McKnight AJ and Gordon S (1998). The EGF-TM7 family: unusual structures at the leukocyte surface. J Leukoc Biol 63, 271–280.
- Perry VH and Gordon S (1991). Macrophages and the nervous system. Int Rev Cytol 125, 203–244.
- Pow DV et al. (1989). Microglia in the neurohypophysis associate with and endocytose terminal portions of neurosecretory neurons. Neuroscience 33, 567–578.
- Qiao T et al. (2023). Dynamic differentiation of F4/80+ tumor-associated macrophage and its role in tumor vascularization in a syngeneic mouse model of colorectal liver metastasis. Cell Death Dis 14, 117.
- Sánchez-Paulete AR et al. (2023). Targeting macrophages with CAR T cells delays solid tumor progression and enhances anti-tumor immunity. Cancer Immunol Res 10, 1354–1369.
- Schaller E et al. (2002). Inactivation of the F4/80 glycoprotein in the mouse germ line. Mol Cell Biol 22, 8035–8043.
- Sprenger-Svačina A et al. (2024). Differential regulation of tissue-resident and blood-derived macrophages in models of autoimmune and traumatic peripheral nerve injury. Front Immunol 15, 1487788.
- Stavrou M and Constantinidou A (2024). Tumor associated macrophages in breast cancer progression: implications and clinical relevance. Front Immunol 15, 1441820.
- Suther T et al. (2003). The brain as an immune privileged site: dendritic cells of the central nervous system inhibit T cell activation. Eur J Immunol 33, 2998–3006.
- Wang J et al. (2017). Tumor cells induced-M2 macrophage favors accumulation of Treg in nasopharyngeal carcinoma. Int J Clin Exp Pathol 10, 8389–8401.
- Wang Y et al. (2001). Blood mononuclear cells induce regulatory NK T thymocytes in anterior chamber-associated immune deviation. J Leukoc Biol 69, 741–746.
- Warschkau H and Kiderlen AF (1999). A monoclonal antibody directed against the murine macrophage surface molecule F4/80 modulates natural immune response to Listeria monocytogenes. J Immunol 163, 3409–3416.
- Wilbanks GA and Streilein JW (1991). Studies on the induction of anterior chamber-associated immune deviation (ACAID). 1. Evidence that an antigen-specific, ACAID-inducing, cell-associated signal exists in the peripheral blood. J Immunol 146, 2610–2617.
- Wilbanks GA et al. (1991). Studies on the induction of anterior chamber-associated immune deviation (ACAID). II. Eye-derived cells participate in generating blood-borne signals that induce ACAID. J Immunol 146, 3018–3024.
- Wilbanks GA et al. (1992). Studies on the induction of anterior chamber-associated immune deviation (ACAID). III. Induction of ACAID depends upon intraocular transforming growth factor-beta. Eur J Immunol 22, 165–173.
- Yagi T et al. (2019). Tumour-associated macrophages are associated with poor prognosis and programmed death ligand 1 expression in oesophageal cancer. Eur J Cancer 111, 38–49.
Be the First to Receive Updates on Products and Resources
You'll receive regular communications featuring free lab resources such as application guides, posters, mini-reviews, protocols, practical tips, webinar invitations, special offers, and updates on new research tools and products.
You'll also get our specialist newsletters—including Fab Fragments, Flow. Sorted., and RiVETing—along with news on the latest antibody innovations, such as TrailBlazer™ Antibodies and StarBright™ Dyes.
During sign-up, you can choose your areas of interest so you only receive content relevant to your research, and you can unsubscribe at any time.
