IgA antibody | K61 1B4
Mouse anti Pig IgA
- Product Type
- Monoclonal Antibody
- Clone
- K61 1B4
- Isotype
- IgG1
- Specificity
- IgA

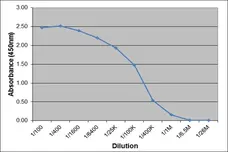
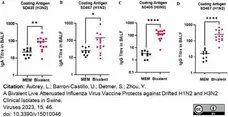
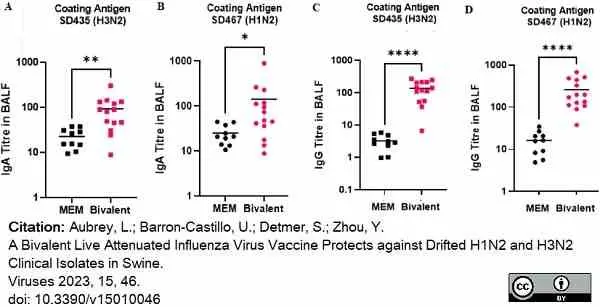
Mouse anti Porcine IgA antibody, clone K61 1B4 (MCA638GA) used to evaluate IgA levels in BALF samples by ELISA.
Image caption:
swIAV-specific antibodies induced after bivalent vaccination in pigs measured in the BALF. Pigs were vaccinated twice with a bivalent vaccine composed of SD191-R342 V (1 × 106 PFU) and SD69-K345 V (1 × 106 PFU) on days 0 and 21. On day 31, the pigs were intratracheally challenged with 1 × 106 PFU of either SD435 or SD467 and then monitored daily for 5 days post-challenge. BALF was collected on day 36 at necropsy. (A) IgA levels in BALF against SD435 (H3N2), (B) IgA levels in BALF against SD467 (H1N2), (C) IgG levels in BALF against SD435 (H3N2), and (D) IgG levels in BALF against SD467 (H1N2). Each sample was conducted in duplicate. Each bar represents the average value from each group tested. Significant differences are denoted by * (p < 0.05), ** (p < 0.01), or **** (p < 0.0001). ns = not significant.
From: Aubrey, L.; Barron-Castillo, U.; Detmer, S.; Zhou, Y.
A Bivalent Live Attenuated Influenza Virus Vaccine Protects against Drifted H1N2 and H3N2 Clinical Isolates in Swine.
Viruses 2023, 15, 46.
doi:10.3390/v15010046
This image is from an open access article distributed under terms of a Creative Commons Attribution License.

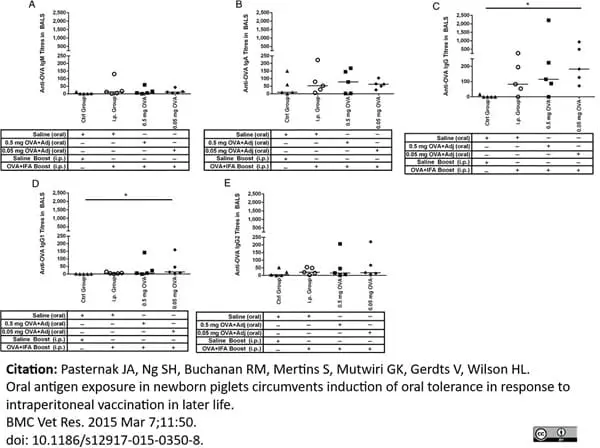
Mouse anti Porcine IgA antibody, clone K61 1B4 (MCA638GA) used for the measurement of ova specific IgA levels in serum by ELISA.
Image caption:
OVA-specific antibody-mediated immune responses in serum from newborn piglets gavaged with OVA then i.p. immunized with OVA at four weeks of age. Piglets (n = 5/group) were gavaged and i.p. immunized as described in Figure 1. Control newborn piglets were not gavaged or immunized with OVA. We measured serum anti-OVA IgM (A), IgA (B), IgG (C), IgG1 (D) and IgG2 (E) production on day 28, day 42 and day 49 after birth. Each data point represents an individual animal and median values are indicated by horizontal lines. *p <0.05., **p <0.01.
From: Pasternak JA, Ng SH, Buchanan RM, Mertins S, Mutwiri GK, Gerdts V, Wilson HL.
Oral antigen exposure in newborn piglets circumvents induction of oral tolerance in response to intraperitoneal vaccination in later life.
BMC Vet Res. 2015 Dec;11(1):350.
This image is from an open access article distributed under the terms of the Creative Commons Attribution License.

Mouse anti Pig IgA antibody, clone K61 1B4 (MCA638) used for the measurement of ova specific IgA levels in lung lavages by ELISA.
Image caption:
OVA-specific antibody-mediated immune responses in lung washes from newborn piglets gavaged with OVA then i.p. immunized with OVA at four weeks of age. Piglets (n = 5/group) were gavaged and i.p. immunized as described in Figure 1. Control newborn piglets were not gavaged or immunized with OVA. Lung lavages were collected four weeks post i.p. immunization and OVA-specific serum IgM (A), IgA (B), IgG (C), IgG1 (D), and IgG2 (E) titres were measured. ELISA titres are expressed as the reciprocal of the highest dilution resulting in a reading of two standard deviations above the negative control. Each data point represents an individual animal and median values are indicated by horizontal lines. *p <0.05, **p <0.01.
From: Pasternak JA, Ng SH, Buchanan RM, Mertins S, Mutwiri GK, Gerdts V, Wilson HL. Oral antigen exposure in newborn piglets circumvents induction of oral tolerance in response to intraperitoneal vaccination in later life.
BMC Vet Res. 2015 Dec;11(1):350.
This image is from an open access article distributed under the terms of the Creative Commons Attribution License.

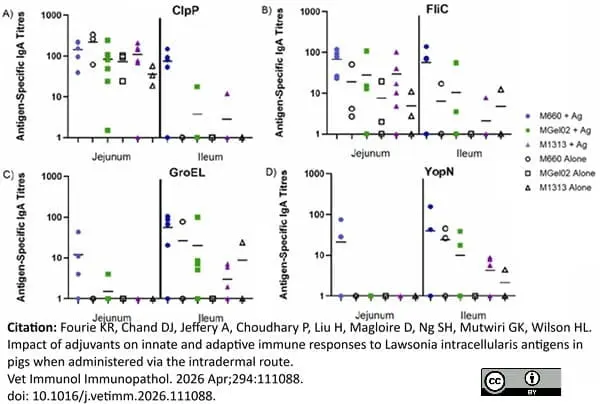
Mouse anti Porcine IgA antibody, clone K61 1B4 (MCA638GA) used to determine antigen specific IgA responsed in pigs by ELISA.
Image caption:
Antigen-specific IgA titres in jejunal and ileal tissues in response to intradermal vaccines with different Montanide adjuvants. No significant antigen-specific IgA titres were observed after intradermal vaccination in response to M660, MGel02 or M1313 adjuvants after 42 days.
From: Fourie KR, Chand DJ, Jeffery A, Choudhary P, Liu H, Magloire D, Ng SH, Mutwiri GK, Wilson HL.
Impact of adjuvants on innate and adaptive immune responses to Lawsonia intracellularis antigens in pigs when administered via the intradermal route.
Vet Immunol Immunopathol. 2026 Apr;294:111088.
doi: 10.1016/j.vetimm.2026.111088.
This image is from an open access article distributed under terms of a Creative Commons Attribution License.
Filter by Application:
E Reset| Mouse Anti Pig IgA antibody, clone K61 1B4 recognizes porcine immunoglobulin (Ig) alpha chain. No cross-reactivity is observed with either porcine IgM or IgG. It has been suggested that pigs possess two subclasses of IgA, referred to as either IgA1 and IgA2 similar to the human IgA subclasses. More recent research indicates that these are allotypic variants and described as IgAa and IgAb (Navarro et al. 2000). Research undertaken in connection with porcine IgA response to Foot and Mouth disease (Pacheco et al. 2010) has demonstrated that clone K61 1B4 recognises both IgAa and IgAb allotypes. IgA is the dominant immunoglobulin found in surface secretions where its role is in the protection of body surfaces. High levels of IgA may be detected in tracheal secretions, saliva, intestinal fluid and urogenital tract secretions. The primary function of IgA is to prevent adherence of bacteria and virus to epithelial surfaces. IgA may also act within epithelial cells where it has a role in interrupting viral replication. |
- Target Species
- Pig
- Product Form
- Purified IgG - liquid
- Preparation
- Purified IgG prepared by affinity chromatography on Protein A from tissue culture supernatant
- Buffer Solution
- Phosphate buffered saline
- Preservative Stabilisers
- 0.09% Sodium Azide (NaN3)
- Carrier Free
- Yes
- Immunogen
- Porcine IgA
- Approx. Protein Concentrations
- IgG concentration 1.0 mg/ml
- Fusion Partners
- Spleen cells from immunised mice were fused with cells of the mouse P3-X63-Ag8.653 myeloma cell line
- Regulatory
- For research purposes only
- Guarantee
- 12 months from date of despatch
This product is shipped at ambient temperature. It is recommended to aliquot and store at -20°C on receipt. When thawed, aliquot the sample as needed. Keep aliquots at 2-8°C for short term use (up to 4 weeks) and store the remaining aliquots at -20°C.
Avoid repeated freezing and thawing as this may denature the antibody. Storage in frost-free freezers is not recommended.
Avoid repeated freezing and thawing as this may denature the antibody. Storage in frost-free freezers is not recommended.
This product has been reported to work in the following applications. This information is derived from testing within our laboratories, peer-reviewed publications or personal communications from the originators. Please refer to references indicated for further information. For general protocol recommendations, please visit the antibody protocols page.
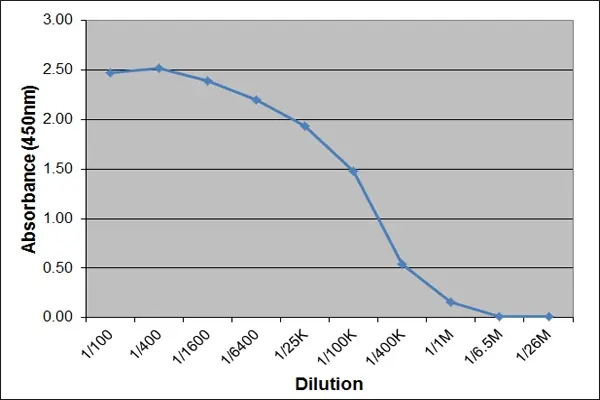
| Application Name | Verified | Min Dilution | Max Dilution |
|---|---|---|---|
| ELISA |  |
1/50K | 1/500K |
| Flow Cytometry | |
||
| Immunohistology - Frozen | |
Where this antibody has not been tested for use in a particular technique this does not necessarily exclude its use in such procedures. Suggested working dilutions are given as a guide only. It is recommended that the user titrates the antibody for use in their own system using appropriate negative/positive controls.
- ELISA
- This product may be used as a detection reagent in ELISA applications
| Description | Product Code | Applications | Pack Size | List Price | Your Price | Quantity | |
|---|---|---|---|---|---|---|---|
| Goat anti Mouse IgG (H/L):HRP (Multi Species Adsorbed) | STAR117P | C E WB | 0.5 mg |

|
Log in | ||
| List Price | Your Price | ||||||
|
|
Log in | ||||||
| Description | Goat anti Mouse IgG (H/L):HRP (Multi Species Adsorbed) | ||||||
| Description | Product Code | Applications | Pack Size | List Price | Your Price | Quantity | |
|---|---|---|---|---|---|---|---|
| Mouse IgG1 Negative Control | MCA928 | F | 100 Tests |
|
Log in | ||
| List Price | Your Price | ||||||
|
|
Log in | ||||||
| Description | Mouse IgG1 Negative Control | ||||||
| Description | Product Code | Applications | Pack Size | List Price | Your Price | Quantity | |
|---|---|---|---|---|---|---|---|
| Mouse anti Pig Ig Lambda Light Chain | MCA633GA | C E F P | 0.1 mg |
|
Log in | ||
| List Price | Your Price | ||||||
|
|
Log in | ||||||
| Description | Mouse anti Pig Ig Lambda Light Chain | ||||||
| Mouse anti Pig IgA Secretory Component | MCA634GA | E | 0.1 mg |
|
Log in | ||
| List Price | Your Price | ||||||
|
|
Log in | ||||||
| Description | Mouse anti Pig IgA Secretory Component | ||||||
| Mouse anti Pig IgG1 | MCA635GA | E | 0.1 mg |
|
Log in | ||
| List Price | Your Price | ||||||
|
|
Log in | ||||||
| Description | Mouse anti Pig IgG1 | ||||||
| Mouse anti Pig IgG2 | MCA636GA | E | 0.1 mg |
|
Log in | ||
| List Price | Your Price | ||||||
|
|
Log in | ||||||
| Description | Mouse anti Pig IgG2 | ||||||
| Mouse anti Pig IgM | MCA637GA | C E F | 0.1 mg |
|
Log in | ||
| List Price | Your Price | ||||||
|
|
Log in | ||||||
| Description | Mouse anti Pig IgM | ||||||
References for IgA antibody
-
Leitão, A. et al. (2001) The non-haemadsorbing African swine fever virus isolate ASFV/NH/P68 provides a model for defining the protective anti-virus immune response.
J Gen Virol. 82 (Pt 3): 513-23. -
Bourges, D. et al. (2004) T and IgA B lymphocytes of the pharyngeal and palatine tonsils: differential expression of adhesion molecules and chemokines.
Scand J Immunol. 60 (4): 338-50. -
Nejsum, P. et al. (2009) Population dynamics of Trichuris suis in trickle-infected pigs.
Parasitology. 136 (6): 691-7. -
Pacheco, J.M. et al. (2010) IgA antibody response of swine to foot-and-mouth disease virus infection and vaccination.
Clin Vaccine Immunol. 17: 550-8. -
Inman CF et al. (2012) Neonatal colonisation expands a specific intestinal antigen-presenting cell subset prior to CD4 T-cell expansion, without altering T-cell repertoire.
PLoS One. 7 (3): e33707. -
Ewaschuk JB et al. (2012) Barley-derived β-glucans increases gut permeability, ex vivo epithelial cell binding toE. coli, and naive T-cell proportions in weanling pigs.
J Anim Sci. 90 (8): 2652-62. -
Kandasamy, S. et al. (2014) Lactobacilli and Bifidobacteria enhance mucosal B cell responses and differentially modulate systemic antibody responses to an oral human rotavirus vaccine in a neonatal gnotobiotic pig disease model.
Gut Microbes. 5 (5): 639-51. -
Kandasamy, S. et al. (2014) Prenatal vitamin A deficiency impairs adaptive immune responses to pentavalent rotavirus vaccine (RotaTeq®) in a neonatal gnotobiotic pig model.
Vaccine. 32 (7): 816-24.
View The Latest Product References
-
Kringel H et al. (2015) Serum antibody responses in pigs trickle-infected with Ascaris and Trichuris: Heritabilities and associations with parasitological findings.
Vet Parasitol. 211 (3-4): 306-11. -
Guzman-Bautista, E.R. et al. (2015) Tracheal and bronchial polymeric immunoglobulin secretory immune system (PISIS) development in a porcine model.
Dev Comp Immunol. 53 (2): 271-82. -
Pasternak, J.A. et al. (2015) Oral antigen exposure in newborn piglets circumvents induction of oral tolerance in response to intraperitoneal vaccination in later life.
BMC Vet Res. 11 (1): 350. -
Makadiya, N. et al. (2016) S1 domain of the porcine epidemic diarrhea virus spike protein as a vaccine antigen.
Virol J. 13: 57. -
Aubrey, L. et al. (2022) A Bivalent Live Attenuated Influenza Virus Vaccine Protects against Drifted H1N2 and H3N2 Clinical Isolates in Swine
Viruses. 15 (1): 46. -
Fourie, K.R. et al. (2024) Vaccination with a Lawsonia intracellularis subunit water in oil emulsion vaccine mitigated some disease parameters but failed to affect shedding.
Vaccine. 42 (24): 126254. -
Zuckermann, F.A. et al. (2024) An effective vaccine against influenza A virus based on the matrix protein 2 (M2)
Veterinary Microbiology. 298: 110245. -
Amimo, J.O. et al. (2024) Maternal immunization and vitamin A sufficiency impact sow primary adaptive immunity and passive protection to nursing piglets against porcine epidemic diarrhea virus infection.
Front Immunol. 15: 1397118. -
Fourie, K.R. et al. (2026) Impact of adjuvants on innate and adaptive immune responses to Lawsonia intracellularis antigens in pigs when administered via the intradermal route.
Vet Immunol Immunopathol. 294: 111088.
Further Reading
-
Navarro, P. et al. (2000) Porcine IgA allotypes are not equally transcribed or expressed in heterozygous swine.
Mol Immunol. 37: 653-64.
MCA638GA
If you cannot find the batch/lot you are looking for please contact our technical support team for assistance.
View more products with IgA specificity
Please Note: All Products are "FOR RESEARCH PURPOSES ONLY"
View all Anti-Pig ProductsAlways be the first to know.
When we launch new products and resources to help you achieve more in the lab.
Yes, sign me up